En esta sección se abordará la biomolécula de principal fuente energética
para nuestro organismo, los carbohidratos. Los carbohidratos son unas biomoléculas que además toman los nombres de
hidratos de carbono, glúcidos, azúcares o sacáridos; aun cuando ambos
primeros nombres, los más frecuentes y empleados, no son enteramente
exactos, debido a que no se tratan estrictamente de átomos de carbono
hidratados, sin embargo los intentos por suplir dichos términos por otros
más exactos no tuvieron triunfo. Estas moléculas permanecen formadas por 3
recursos primordiales: el carbono, el hidrógeno y el oxígeno, este último en
una proporción algo más baja. Su primordial funcionalidad en el organismo de
los organismos vivos es la de contribuir en el almacenamiento y en la
obtención de energía de manera instantánea, más que nada al cerebro y al
sistema nervioso.
Función de los carbohidratos en nuestro organismo
Los carbohidratos son la primordial fuente de energía del organismo ya que
a lo largo de su digestión se produce glucosa, siendo este el combustible
querido por las células del organismo, debido a que degradan esta molécula y
se crea ATP, el cual es usado en los múltiples procesos metabólicos para el
buen desempeño del organismo. La glucosa es usada primordialmente por el
cerebro, usando 120 gramo por día, lo cual representa la mayor parte de los
160 gramo de glucosa que el cuerpo humano requiere diariamente.
Además de esto, una sección de la glucosa generada es almacenada a modo de
glucógeno en el hígado, y una pequeña cantidad en los músculos, gracias a
probables eventualidades en las que el organismo ocupe usar esta reserva
como puede pasar en situaciones de ayuno prolongado, de alerta o estrés
metabólico, ejemplificando.
El consumo de carbohidratos además es fundamental para la preservación de
los músculos, debido a que la carencia de glucosa favorecerá la pérdida de
masa muscular. Asimismo, la fibra además es un tipo de carbohidrato, que
pese a que no es digerida en glucosa, es sustancial para el proceso de
digestión, debido a que reduce la absorción de colesterol, ayuda a conservar
el azúcar en la sangre, se incrementa los movimientos del intestino y
beneficia el incremento del volumen de las heces, evitando inconvenientes
como el estreñimiento.
Clasificación
Simples
Los carbohidratos simples o monosacáridos son unidades o moléculas básicas
que al unirse conforman carbohidratos más complicados, dichos son la
glucosa, la ribosa, la xilosa, la galactosa y la fructosa. Al consumir una
cantidad de carbohidrato, esta molécula más compleja se irá descomponiendo a
grado del tracto digestivo hasta llegar al intestino a modo de monosacáridos
para lograr ser absorbidos.
La unión de dos unidades de monosacáridos forman disacáridos como la
sacarosa o el azúcar de mesa (glucosa + fructosa), la lactosa (galactosa +
glucosa) y la maltosa (glucosa + glucosa), por ejemplo. Además de esto, la
unión de 3 a 10 unidades de monosacáridos dan origen a oligosacáridos.
Complejos
Los carbohidratos complejos o polisacáridos son aquellos que contienen más
de 10 unidades de monosacáridos, formando estructuras moleculares
complejas que pueden ser lineares o ramificadas, algunos ejemplos son el
almidón, el glucógeno que se almacena en el hígado y la celulosa.
Dentro de los carbohidratos complejos se encuentra la fibra, que son
componentes de los vegetales que no son digeridos por las enzimas
gastrointestinales, algunos ejemplos son la celulosa, los
fructooligosacáridos (FOS) y la lignina.
Digestión y absorción de carbohidratos
La digestión del almidón se inicia en la boca, durante la masticación, ya
que en la saliva se encuentra una hidrolasa, que recibe el nombre de
amilasa salival, la cual, introduciendo una molécula de agua, rompe el
enlace glucosídico α - 1 —> 4, que mantiene unidas a las moléculas de
glucosa en el polímero. Cada vez que actúa la enzima se produce una
molécula de glucosa libre y almidón, que tiene una unidad menos de las que
tenía en un principio.
La acción de la amilasa salival dura únicamente mientras los alimentos
pasan de la boca hacia el estómago, a través del esófago, debido a que el
pH del estómago es muy bajo y el pH óptimo de la amilasa salival es
cercano a 7. Por ello la amilasa salival se inactiva al llegar a este
órgano.
En el estómago los carbohidratos no sufren ninguna transformación
química.
Es en el intestino delgado en donde ocurre la mayor parte de la digestión
de los carbohidratos, ya que ahí se secretan los fluidos producidos por el
páncreas y algunas células de las paredes del intestino, que llevan en
solución enzimas específicas para hidrolizar carbohidratos.
El páncreas sintetiza la amilasa pancreática, que actúa de manera
idéntica a la salival, pero durante el tiempo suficiente para lograr la
degradación total de una molécula de almidón hasta glucosa. Las dos
amilasas que se han analizado rompen solamente enlaces glucosídicos α - 1
—> 4. En el caso de la amilopectina que tiene ramificaciones α - 1
—> 6, se requiere además otra enzima, producida también por el
páncreas, que hidroliza estos enlaces para lograr su degradación total
hasta glucosa.
En el hígado los monosacáridos diferentes a la glucosa son convertidos a
este compuesto; la glucosa "nueva" puede seguir dos rutas: ser liberada a
la sangre para ser transportada hacia otros tejidos del organismo, o ser
almacenada en forma de glucógeno, constituyendo así una reserva de
carbonos y de energía que será usada cuando el organismo lo demande y en
esos momentos no haya otra fuente de energía disponible
Rutas metabólicas de los carbohidratos
Glucolisis
La glucolisis tiene lugar en el citoplasma celular. Consiste en una
serie de diez reacciones, cada una catalizada por una enzima
determinada, que permite transformar una molécula de glucosa en dos
moléculas de un compuesto de tres carbonos, el ácido pirúvico.
En la primera parte se necesita energía, que es suministrada por dos
moléculas de ATP, que servirán para fosforilar la glucosa y la fructosa.
Al final de esta fase se obtienen, en la práctica dos moléculas de PGAL,
ya que la molécula de DHAP (dihidroxiacetona-fosfato), se transforma en
PGAL.
En la segunda fase, que afecta a las dos moléculas de PGAL, se forman
cuatro moléculas de ATP y dos moléculas de NADH. Se produce una ganancia
neta de dos moléculas de ATP.
Al final del proceso la molécula de glucosa queda transformada en dos
moléculas de ácido pirúvico, es en estas moléculas donde se encuentra en
estos momentos la mayor parte de la energía contenida en la glucosa.
La glucolisis se produce en la mayoría de las células vivas, tanto en
procariotas como en las eucariotas.
Glucogenólisis
El glucógeno es un polímero de glucosa
unidas por dos tipos de enlaces: α(1,4)
y α(1,6) glucosídicos. Se encuentra en el citosol, y
sus principales lugares de almacenamiento son el hígado y el músculo
esquelético.
Es una macromolécula de una estructura
muy ramificada y en su núcleo
existe una proteína central: la glicogenina (una
glicoproteína extraordinariamente glicosilisada)
La glucógeno fosforilasa rompe los enlace glucosídicos α(1,4) entre los
residuos que están en los extremos no reductores por simple fosforilación.
La enzima que cataliza este proceso contiene al fosfato piroxidal unido
covalentemente como coenzima. Es una
fosfotranferasa que degrada secuencialmente
las cadenas de glucógeno en sus
extremos no reductores hasta que
quedan sólo cuatro residuos de glucosa
en la ramificación. Las ramas de glucógeno son removidas por medio
de dos actividades enzimáticas.
Primero la glucosil transferasa remueve tres de los cuatro residuos unidos
a la rama y los transfiere a un extremo no reductor de otra rama. Rompe un
enlace alfa (1,4) pero forma otro
A continuación el residuo restante de
glucosa unido a la cadena en
posición α(1,6), es removido hidrolíticamente por la
amilo-α-(1,6) glucosidasa liberando glucosa. La cadena glucosídica
es ahora accesible a la degradación por la glucógeno fosforilasa. La
glucosa-1-fosfato es trasformada en glucosa-6-fosfato por la acción de la
enzima fosfoglucomutasa. En el músculo, la glucosa-6-P es metabolizada por
las enzimas de la glucólisis (piruvato), generando energía para su propio
consumo. En el hígado, la glucosa-6-P además es convertida en glucosa por
acción de la enzima glucosa-6-fosfatasa, ausente en el músculo es por esta
razón que no aportan glucosa a la sangre), y liberada

Glucogenogenesis
La glucogenogénesis es la ruta
anabólica por la que tiene lugar
la síntesis de glucógeno a partir
de un precursor más simple: la glucosa-6-fosfato. Se lleva a cabo
principalmente por el hígado, y es activado por la insulina en respuesta a
los altos niveles de glucosa, que pueden ser por ejemplo posteriores a la
ingesta de alimentos con carbohidratos.
Se produce gracias a la enzima glucógeno sintasa. La adición de una
molécula de glucosa al glucógeno consume dos enlaces de alta
energía: una procedente del ATP y otra que procede del UTP.
La síntesis del glucógeno tiene lugar en varios pasos:
En primer lugar, la glucosa es transformada en glucosa-6-fosfato,
gastando una molécula de ATP.
glucosa + ATP → glucosa-6-P + ADP
A continuación se transforma la glucosa-6-fosfato en
glucosa-1-fosfato
glucosa-6-P ←→ glucosa-1-P
Se transforma la glucosa-1-fosfato en UDP-glucosa, con el gasto de
un UTP.
glucosa-1-P + UTP → UDP-glucosa + PPi
La glucógeno sintasa (con acción antagónica a la glucógeno
fosforilasa), que no gasta ATP, va uniendo UDP-glucosa para formar
el glucógeno, mediante enlaces alfa 1-4 liberando el nucleótido UDP
(que se reutilizará).
(glucosa)n + UDP-glucosa → (glucosa)n+1 + UDP
La enzima ramificadora del glucógeno se encarga de ramificar la
cadena introduciendo enlaces glucosídicos alfa 1-6.
Puesto que la glucógeno sintasa necesita una cadena preexistente
para empezar su acción, hay otra enzima que se encarga de catalizar
el comienzo de la síntesis del glucógeno: la glucogenina, capaz de
crear un enlace covalente sobre un grupo hidroxilo (-OH) de un
residuo de tirosina (Tyr) de su propia molécula y fijar la primera
glucosa de la cadena; acto seguido podrá actuar la glucógeno sintasa
y una vez añadidos unos 10-12 residuos de glucosa la glucogenina
dejará de ser imprescindible, separándose y dejando espacio para las
ramificaciones siguientes.
Ciclo de pentosas fosfato
El ciclo de la pentosa fosfato es una ruta metabólica en la que, por
una parte, se obtiene poder reductor en forma de NADPH y, por otra
parte, se producen azúcares muy importantes para la síntesis de ácidos
nucleicos.
Todo empieza con una molécula de glucosa. Esta glucosa suele entrar en
una vía catabólica conocida como glucólisis que se basa en degradarla
para obtener energía, pero también puede entrar en este ciclo de la
pentosa fosfato. A partir de aquí, se entra en la ruta metabólica, que
se divide en dos partes: la fase oxidativa y la no oxidativa.
La primera de las fases es la oxidativa y es en la que se genera todo
el NADPH de la ruta. En esta fase, la glucosa se convierte primero en
glucosa 6-fosfato, la cual, a través de la enzima más importante del
ciclo (la glucosa-6-fosfato deshidrogenasa), se convierte en otro
metabolito intermedio. Lo importante es que como “efecto colateral” de
la conversión, se libera el NADPH.
A través de otras enzimas se llega hasta la ribulosa-5-fosfato, que
marca el final de la fase oxidativa. En este momento, se ha obtenido
ya todo el NADPH. Pero en caso de que la célula necesite azúcares para
sintetizar ácidos nucleicos, se entra en la fase no oxidativa.
La fase no oxidativa del ciclo de la pentosa fosfato consiste en la
conversión de esta ribulosa-5-fosfato en la ribosa 5-fosfato, un
azúcar que es pieza clave en la síntesis de nucleótidos, las unidades
que constituyen el ADN.
Además, a partir de esta ribosa 5-fosfato y siguiendo con la fase no
oxidativa del ciclo, se pueden sintetizar muchos azúcares distintos
que actúan como metabolitos iniciales (precursores) o intermediarios
de otras rutas, ya sean anabólicas o catabólicas, siendo las pentosas
los más importantes
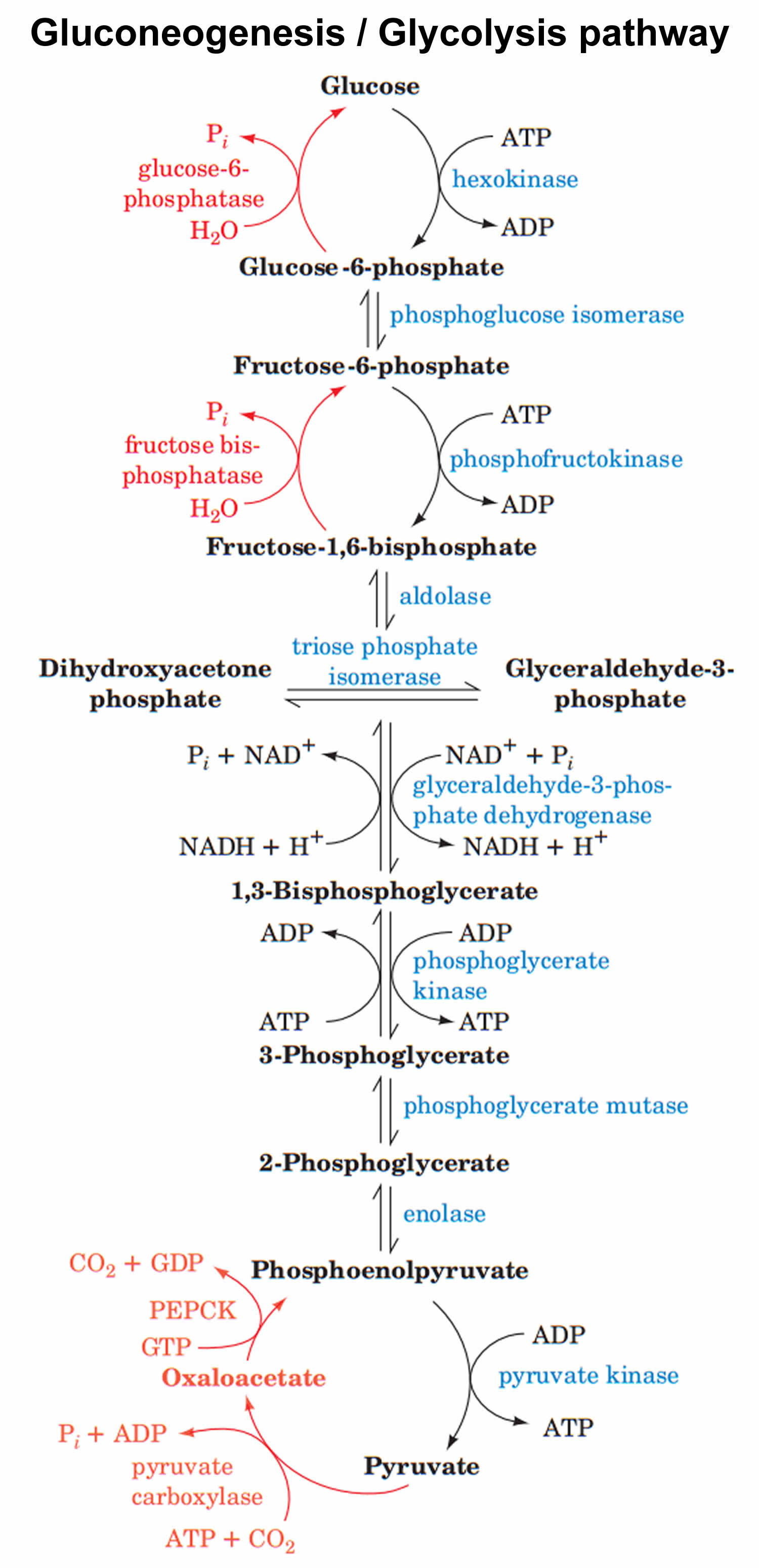
Gluconeogenesis
La gluconeogénesis es una ruta metabólica anabólica que posibilita la
biosíntesis de glucosa desde precursores no glucídicos. Incluye la
implementación de diversos aminoácidos, lactato, piruvato, glicerol y
cualquier persona de los intermediarios del periodo de los ácidos
tricarboxílicos (o periodo de Krebs) como fuentes de carbono para la
vía metabólica. Todos los aminoácidos, excepto la leucina y la lisina,
tienen la posibilidad de proporcionar carbono para la síntesis de
glucosa.
Los Ácidos grasos de cadena par no dan carbonos para la síntesis de
glucosa, puesto que el resultado de su β-oxidación (Acetil-CoA) no es
un sustrato gluconeogénico; en lo que los ácidos grasos de cadena
impar proporcionarán un esqueleto de carbonos que derivarán en
Acetil-CoA y Succinil-CoA (que sí es un sustrato gluconeogénico por
ser un intermediario del periodo de Krebs). Ciertos tejidos, como el
cerebro, los eritrocitos, el riñón, la córnea del ojo y el músculo,
una vez que la persona ejecuta actividad extenuante, necesitan de un
aporte constante de glucosa, obteniéndola desde el glucógeno
perteneciente del hígado, el cual solo puede saciar estas necesidades
a lo largo de 10 a 18 horas como más alto, lo cual tarda en agotarse
el glucógeno guardado en el hígado. Más adelante empieza la formación
de glucosa desde sustratos diferentes al glucógeno.
La gluconeogénesis tiene lugar casi exclusivamente en hígado (10% en
los riñones). Es un proceso clave pues permite a los organismos
superiores obtener glucosa en estados metabólicos como el ayuno.
Las enzimas que participan en la vía glucolítica participan también
en la gluconeogénesis; ambas rutas se diferencian por tres
reacciones irreversibles que utilizan enzimas específicas de este
proceso y los dos rodeos metabólicos de esta vía.
Estas reacciones son:
- De glucosa-6-fosfato a glucosa.
- De fructosa-1,6-bisfosfato a fructosa-6-fosfato.
- De piruvato a fosfoenolpiruvato.











Comentarios
Publicar un comentario